Voluntary Wheel-Running by Ovariectomized Rats: Impacts on Weight Gain, Plasma Leptin and Insulin, Hindbrain Astrocytes and Toll-like Receptor 4
Steven Rivera, M.S III, Oklahoma State University College of Osteopathic Medicine
Donald Wu, M.S III, Oklahoma State University College of Osteopathic Medicine
Randall L. Davis, Ph.D., Department of Pharmacology and Physiology, OSU Center for Health Sciences
Kathleen S. Curtis, Ph.D. Department of Pharmacology and Physiology, OSU Center for Health Sciences
Abstract
Although obesity is an ongoing healthcare challenge in the U.S., diet and exercise have limited efficacy in losing weight and maintaining the weight loss, particularly for post-menopausal women. Nonetheless, few preclinical studies focus on females, even though it is well-known that rats rapidly gain weight after ovariectomy. To evaluate the effect of exercise on weight gain, we gave one group of ovariectomized (OVX) rats access to running wheels for three weeks immediately after ovariectomy; a second group had access to running wheels that was delayed for three weeks after ovariectomy. We also investigated the effect of exercise on circulating metabolic hormones that communicate with the central nervous system (CNS), as well as on neuroimmune factors in CNS areas involved in the control of body weight. Body weight was assessed each week for six weeks, and circulating levels of metabolic hormones and central neuroimmune factors were assessed after six weeks. Regardless of whether exercise was initiated immediately or was delayed for three weeks, OVX rats gained weight at a slower rate during the first week of running. However, this transient effect was longer-lasting when running was delayed. Circulating insulin and leptin were decreased in OVX rats when running was delayed; hindbrain astrocyte activation and toll-like receptor 4 levels also were reduced. Thus, although adaptations to voluntary exercise may limit its efficacy as a weight loss strategy, even transient slowing of post-ovariectomy weight gain is associated with reductions in metabolic hormones, which may be related to region-specific modulation of central neuroimmune factors.
Introduction
In the United States, ~40% of adults are obese1,2, with ~7% classified as morbidly obese3. Obesity is associated with a range of co-morbidities, including type 2 diabetes, stroke, heart disease, kidney disease, and cancer1,3. The associated medical costs are staggering: people with obesity are estimated to pay as much as $2,700 more in health care costs annually compared to normal weight individuals4. Managing obesity remains a challenge, in part, because diet and exercise, the most common recommendations for obese individuals1, typically result in only short-lived weight loss5,6. Further complicating the issue is that obesity is a multi-faceted disease that is influenced by genetics and the environment. For example, obesity disproportionately affects African Americans, Hispanics, and groups with lower education and socioeconomic status1,2. Moreover, more women are obese compared to men (38.0% vs. 41.5%2), with a further increase in obesity in post-menopausal women2,7. The increase in obesity after menopause is particularly troubling given the aging of the U.S. population. Surprisingly, obesity research has largely focused on males and this sex bias extends to preclinical studies involving animal models. Most of these studies preferentially use younger animals, further limiting their applicability to understanding and managing obesity in post-menopausal women.
Female rats rapidly and reliably gain weight after ovariectomy8,9 and the weight gain follows a predictable time course10. A variety of mechanisms likely contributes to this body weight gain11, among which are peripheral signals associated with eating and body weight including metabolic hormones such as leptin and insulin11. These hormones have actions at receptors localized to the hypothalamic arcuate (ARC) and paraventricular (PVN) nuclei12-13, and the hindbrain dorsal vagal complex (DVC), central nervous system (CNS) areas that studies of male rodents have shown to be involved in body weight regulation and control of feeding14-16. In our recent study, we found that weight gain in OVX rats is associated with changes in chemokine and cytokine expression in the ARC, PVN, and DVC10, suggesting that reproductive hormones also alter central neuroimmune mechanisms that impact body weight regulation. Notably, these changes in neuroimmune signals followed a temporal pattern, with some occurring early during the post-ovariectomy weight gain, and others not evident until the rate of weight gain had slowed.
Taken together, these findings raise the possibility that strategies like exercise, commonly used to manage body weight, may be differentially effective depending on the time at which the strategies are initiated, and on their impact on metabolic hormones which, in turn, may affect neuroimmune signals. Accordingly, in the current studies, we introduced voluntary exercise at different times after ovariectomy to evaluate the impact on post-ovariectomy weight gain and on peripheral metabolic hormone levels. At the same time, we conducted preliminary assessments of central neuroimmune factors to determine whether central neuroimmune signals in ovariectomized (OVX) rats are modified by voluntary running and/or changes in metabolic hormones, and whether the timing of initiating exercise alters these effects.
Materials and Methods
Animals
Adult female Sprague-Dawley rats (90 days; Charles River) were individually housed in a temperature-controlled room (21-25℃) in animal facilities at the Oklahoma State University-Center for Health Sciences (OSU-CHS) on a 12:12 light/dark cycle (lights on at 07:00), with unlimited access to standard rodent chow and water. All procedures were approved by the OSU-CHS Animal Care and Use Committee and were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Experimental Procedures
After seven days of acclimation to the colony room, rats (n=16) were treated with the long-lasting analgesic, meloxicam (1.5 mg/kg, i.o.) before anesthesia with isoflurane. Bupivacaine was applied topically, the abdomen was incised, the ovaries were removed, and muscle and skin then were closed; ~18 h later, bupivacaine was applied to the incision site. Rats were allowed to recover for an additional 24 h. Body weights of rats were recorded immediately prior to ovariectomy, and then at weekly intervals for a total of six weeks. Change in body weight was calculated each week as [weight (g) for current week - weight (g) for previous week].
During the first three weeks, one group of rats (n=8) was individually housed in standard cages (44 × 22 × 20 cm). The other group (n=8) had 24 h/day access to 35 cm-diameter running wheels (Lafayette Instruments) attached to individual plexiglass cages (47 × 37 × 20 cm). Running distance data were collected using electronic counters linked to a personal computer with Lafayette Instruments software.
After the first three weeks, the groups were switched, and rats that initially had access to running wheels were housed in standard cages (R-S), while rats that initially were housed in standard cages then were housed in cages with attached running wheels (S-R). This phase lasted another three weeks. Thus, all rats had three weeks of voluntary exercise, with the R-S group given access to running wheels immediately after ovariectomy and the S-R group given access beginning three weeks after ovariectomy.
Tissue Collection and Preparation
After 6 weeks, all rats were rendered unconscious by CO2 inhalation and rapidly decapitated. Trunk blood was collected on ice, then centrifuged. Plasma was collected and stored at -80℃ until assayed for total protein concentration and levels of insulin and leptin. Brains and spleens were collected on ice. Brains were stored at -80℃ until processed for determination of neuroimmune factors.
Peripheral factors
Spleens from rats in the R-S group (n=8) and the S-R group (n=8) were weighed; spleen weight was expressed as mg/100 g body weight. Plasma was used to assess circulating levels of the metabolic hormones insulin and leptin, as well as plasma protein concentration as an index of plasma volume.
Insulin and Leptin. Circulating insulin and leptin were assessed using commercially available ELISA kits (Millipore/Sigma; Rat/Mouse Insulin; EZRMI-13K and Rat Leptin; EZRL-83K) according to the manufacturer's instructions.
Plasma protein concentration. Plasma protein concentrations were assessed using a refractometer (Leica).
Neuroimmune factors
ARC, PVN, and DVC were identified in thick, frozen brain sections using the following landmarks: ARC-caudal 3rd ventricle and median eminence; PVN-rostral 3rd ventricle, anterior commissure, and optic chiasm; DVC-central canal, pyramidal tract, and caudal inferior olivary nucleus. Bilateral punches were taken from these areas using a 1.0 mm Uni-Core punch (Ted Pella), collected into Eppendorf tubes, and stored at -80 ℃ until homogenized. Brain tissue (100 mg/ml lysis buffer) was sonicated on ice for 10 sec. in lysis buffer containing 50 nM Tris-HCl, pH 7.4, 150 nM NaCl, 0.02% sodium azide, 0.1% sodium dodecyl sulfate, 1.0% IGEPAL, 0.5% sodium deoxycholate, 10 µg/ml aprotinin and 0.5 mM phenylmethylsulfonyl fluoride.
In these studies, we focused on interleukin-6 (IL-6), monocyte chemoattractant protein-1 (MCP-1), toll-like receptor 4 (TLR4), and glial fibrillary acidic protein (GFAP; a marker of astrocyte activation) because of their involvement in innate immune activation associated with obesity17,18, and the changes in these factors observed in our recent study10.
IL-6 and MCP-1 expression. Homogenized samples of ARC, PVN, and DVC from rats in the R-S group (n=4) and rats in the S-R group (n=4) were assessed for IL-6 and MCP-1 expression by ELISA using commercially available, standard dual-antibody solid phase immunoassay kits (Peprotech, Rocky Hill, NJ) via colorimetric analysis.
GFAP and TLR4 expression. Homogenized samples of ARC, PVN, and DVC from other rats in the R-S group (n=4) and other rats in the S-R group (n=4) were assessed for GFAP and TLR4 by western blot using antibodies from Cell Signaling (Danvers, MA), Santa Cruz Biotechnology (Dallas, TX), MilliporeSigma (Burlington, MA) or Sigma-Aldrich (St. Louis, MO) and standard protocols. Equal amounts of total protein (30 ug) were separated by SDS-PAGE and transferred to PVDF membrane which was incubated in target-specific 1° antibody (anti-TLR4, 1:200; anti-GFAP, 1:5000; orβ -tubulin, 1:1000), washed, then incubated in 2° antibody (anti-IgG, 1:1000). For subsequent targets, membranes were stripped of antibody by western blot stripping buffer (Thermo Fisher) for 20 min at room temperature, washed three times in Tris-buffered saline+Tween 20 (TBST), then re-blocked in 5% bovine serum albumin in TBST for 2 h prior to incubation with 1°/2° antibodies as described above. Labeled proteins were analyzed by NIH Image J software and expressed relative to β-tubulin.
Statistics
All data are shown as group means ± standard error of the mean (SE). Data for body weight, change in body weight, and distance run were analyzed using repeated measures analysis of variance (rmANOVA), with group (R-S, S-R) and week as factors, repeated for week. Significant (p<0.05) main effects or interactions were further assessed using Fisher's LSD tests. Effect sizes were calculated using partial eta squared (pȠ2). Data for peripheral hormone levels, plasma protein concentrations, and spleen weights were assessed using individual 2-tailed t-tests. Effect sizes for these comparisons were calculated using Cohen's d. Data for neuroimmune factors showed considerable variability. Thus, we included only individual data points that were < 2 standard deviations from the group mean for each area prior to analyses using individual Mann-Whitney U tests for nonparametric comparisons.
Results
Body weight
As expected, OVX rats rapidly gained weight (Figure 1A); however, rmANOVA revealed differences in body weight that depended on the interaction between group and week [F(6,84) = 10.48, p<0.001; pȠ2 = 0.428]. Pairwise comparisons of the interaction showed that body weight of OVX rats that ran initially and then were switched to sedentary conditions (R-S) increased steadily, with weight each week significantly greater than each of the preceding weeks (ps<0.05-0.001). In contrast, body weight of OVX rats that initially were sedentary increased only until they began running. For rats in this S-R group, weight during each of the first three weeks was significantly greater than each of the preceding weeks (ps<0.001). After the switch, body weight did not differ from week-to-week; however, weight on week 6 was significantly greater than that on week 4 (p<0.01).
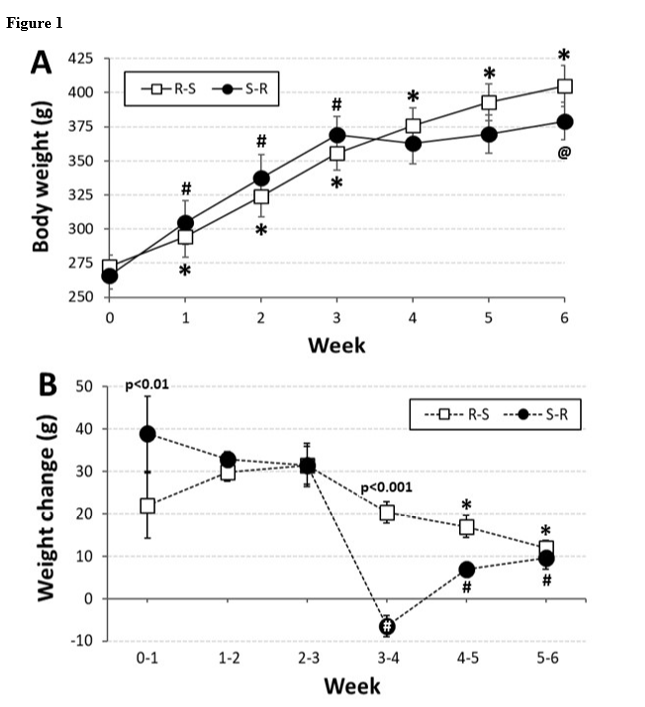
Evaluation of the change in body weight from week to week (Figure 1B) also revealed differences that depended on the interaction between group and week [F(5,70) = 5.21, p<0.001; pȠ2 = 0.271]. Pairwise comparisons of this interaction showed that the R-S group, which ran immediately after ovariectomy, had a consistent weight gain each week, though the gain during the 2nd and 3rd week (while running) both were greater than those during the last 2 weeks (while sedentary; ps<0.05-0.01). The S-R group, which ran after a 3-week delay, also had a consistent weight gain during the first 3 weeks (while sedentary); however, they lost weight during the first week after the switch (while running; ps<0.001, 0.05 vs. all other weeks). For this group, the weight gain during the last 2 weeks were significantly less than the first 3 weeks, but were not different from each other. Finally, during the first week, the R-S group gained significantly less weight (p<0.01), than did the S-R group, but during the week after the switch, the S-R group gained less weight than did the R-S group (p<0.001). There were no other differences between the groups on a weekly basis.
Running distance
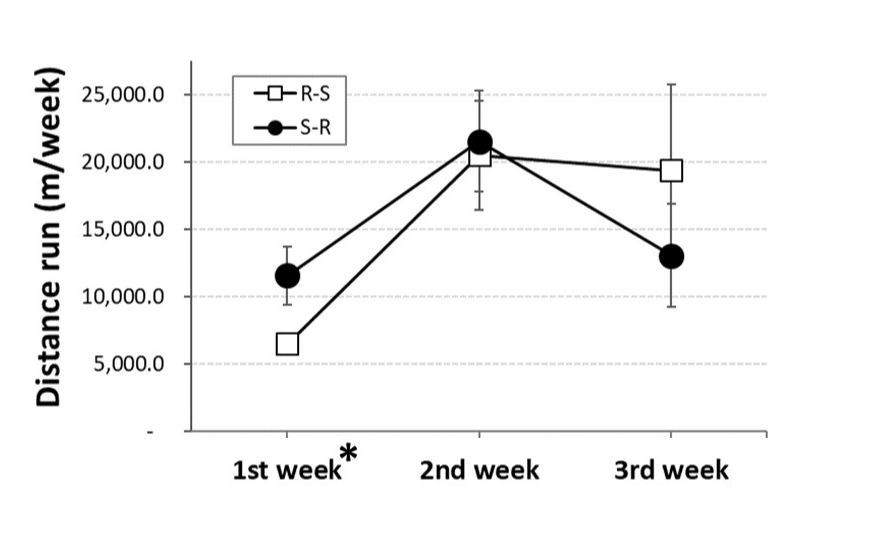
The distance run by OVX rats was not affected by whether they were given access to running wheels immediately or 3 weeks after ovariectomy (Figure 2). Rather, the distance run depended on week [F(2,12) = 9.53, p<0.01; pȠ2 = 0.614]. Pairwise comparisons revealed that, independent of group, OVX rats ran significantly less during their first week of access to running wheels compared to the next two weeks (ps<0.05, 0.001), which were not different from each other.
Figure 2
Peripheral effects
Running condition did not affect spleen weight (Table 1). However, t-tests revealed that plasma protein concentration was significantly less in the S-R group that initially were sedentary and then had access to running wheels for three weeks (Table 1; t(14) = 2.49, p<0.05; d = 0.522).
Table 1

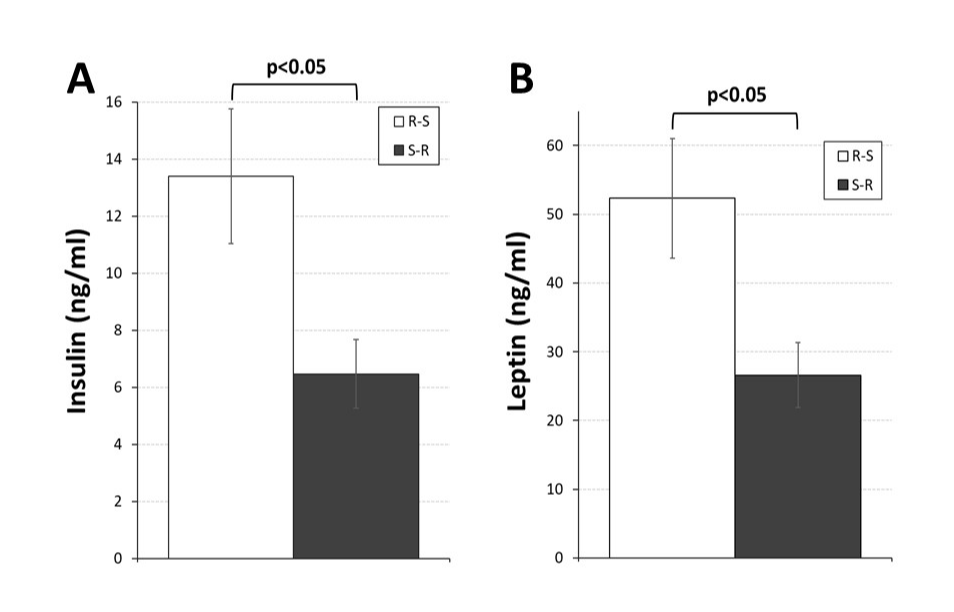
Moreover, both circulating insulin and leptin levels (Figure 3) were significantly less (t(14) = 2.61, p<0.05, d = 0.547; t(14) = 2.67, p<0.05, d = 0.544, respectively) in OVX rats in the S-R group.
Figure 3
Neuroimmune factors
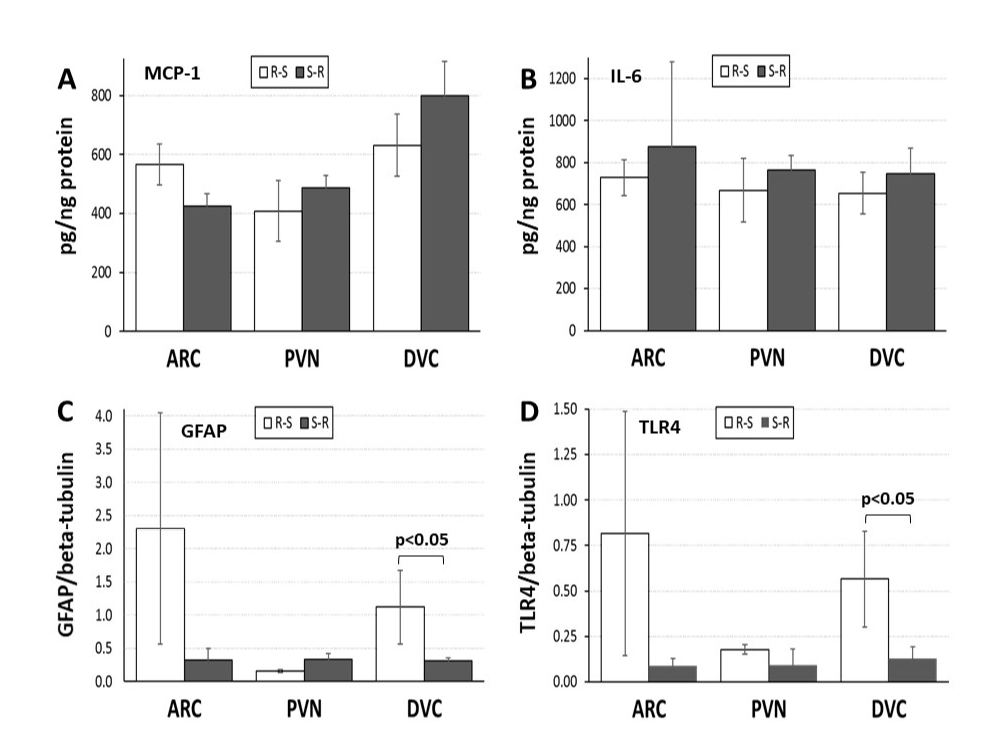
Individual comparisons of neuroimmune factors in the ARC, PVN, and DVC (Figure 4A-D) using Mann-Whitney nonparametric tests revealed that both GFAP and TLR4 levels in the DVC were significantly less in the S-R group that initially were sedentary and then had access to running wheels for three weeks (Z = 2.309, p<0.05; Z = 2.021, p<0.05, respectively). No differences in MCP-1 or IL-6 levels were observed in the DVC, nor did any neuroimmune factors differ in ARC or PVN.
Figure 4
Discussion
The peripheral signals and central pathways involved in the post-ovariectomy weight gain in female rats8,9 remain a subject of more than theoretical interest given the obesity epidemic in the United States1-3,5. Importantly, the weight gain in OVX rats follows a predictable time course10 that allows for systematic evaluation of the impact of weight loss strategies such as diet and exercise-which typically are recommended as the first line of defense1-on body weight, peripheral signals, and central factors implicated in body weight regulation. To date, most investigations have focused on central neurotransmitter systems14-16; however, we recently reported that post-ovariectomy weight gain also is associated with specific temporal patterns of changes in neuroimmune factors10. Accordingly, in the present study, we initiated voluntary exercise at different times after ovariectomy and assessed weight gain and peripheral hormone levels, as well as the impact of exercise on neuroimmune factors in OVX rats.
We found that exercise had transient effects to slow post-ovariectomy weight gain, regardless of whether it was initiated immediately after ovariectomy or was delayed for three weeks. This effect occurred when the distance run was least, raising the possibility that after the first week, some adaptation to increased energy expenditure associated with running19 occurred that minimized weight loss as time went on, even as running increased. Interestingly, when access to running wheels was delayed for three weeks after ovariectomy, rats lost weight during the first week of running and gained more slowly thereafter. Our previous work showed that post-ovariectomy weight gain slows after three weeks10; thus, it is possible that the weight loss observed when access to running wheels was delayed for three weeks was attributable to slower rate of weight gain during this period. However, two observations argue against this interpretation. First, the difference in weight change between runners and sedentary OVX rats was greater when running was delayed vs. when rats ran immediately after OVX (~27 g vs. ~17 g, respectively). Second, despite the 'rebound' in weight gain after the first week of running, it took longer for OVX rats to achieve the same rate of weight gain as their sedentary counterparts when access to running wheels was delayed.
The regulation of body weight incorporates both short-term and long-term controls14-16,20 including signals from the GI tract related to feeding and energy stores, as well as metabolic hormones and other circulating factors. Levels of both insulin and leptin were decreased in rats that initially were sedentary and then had access to running wheels during the last three weeksand see 21. On the surface, it seems reasonable to assume that loss of body weight during the first week of running included reduced white adipose tissue, the source of leptin22, while the blunted insulin levels indicate more efficient glucose handling40, or decreased food intake and/or increased water intake23, as suggested by lower plasma protein concentrations in rats that initially were sedentary and then had access to running wheels. However, leptin and insulin levels were reduced at a time when group differences in body weight no longer were apparent. It is possible that changes in these hormones reflect physiological changes due to increased energy expenditure during running19. Alternatively, the rebound in body weight during the three weeks that R-S rats were sedentary produced changes (e.g. increased adipose, alterations in feeding and/or metabolic rate) leading to increased leptin and insulin in that group. More experiments will be necessary to distinguish between these possibilities; nonetheless, the differences were unexpected given the lack of group differences in body weight at this time point, and lead to the question of how increased body weight is sustained at similar levels in OVX rats despite access to running wheels and differences in insulin and leptin levels.
Although much obesity research focuses on neurotransmitters in CNS pathways14-16, increasing experimental attention has turned to the role of inflammation. The metabolic endotoxemia hypothesis of obesity postulates the involvement of low-grade inflammation associated with activation of TLR4 by classic proinflammatory factors like lipopolysaccharide and/or by free fatty acids, changes that have been shown to occur in rodents during chronic maintenance on a high fat dietfor reviews, see 17,18,25. In fact, much of the evidence linking inflammation and obesity derives from studies that utilize high fat diet-induced obesity (DIO), many of which address peripheral inflammatory effects, such as insulin resistance25. However, recent advances indicate a role for neuroinflammation in DIO. For example, activation of TLR4 on microglia in the hypothalamus may contribute to DIO25 but see 26. To date, however, most investigators have focused on neuroimmune factors in the pathological consequences of extreme and established obesity after prolonged maintenance on high fat diet, with few investigations focused on more physiological weight gain such as that after ovariectomy.
We previously reported that neuroimmune factors in the ARC, PVN, and DVC of OVX rats change as early as three weeks after ovariectomy10. Surprisingly, then, there were no differences between immediate and delayed exercise groups for most neuroimmune factors in the current study. It is possible that the sensitivity of the analytical kits were insufficient to reveal subtle changes in these factors, particularly given that weight did increase in both groups-albeit at slower rates in OVX rats that had access to running wheels during the last three weeks. However, differences in neuroimmune factors were detected in the DVC. Specifically, levels of both GFAP and TLR4 were significantly decreased in the DVC of OVX rats that initially were sedentary and then had access to running wheels for three weeks. In contrast, OVX rats that initially had access to running wheels but were sedentary during the final three weeks exhibited levels of GFAP and TLR4 in the DVC that were comparable to levels observed in OVX rats that did not exercise reported in our previous study. Direct comparisons between the two studies must be interpreted cautiously due to differences in the lots of the analytic kits; nonetheless, together these data suggest that voluntary exercise had selective effects on neuroimmune factors in the DVC.
The contribution of neuroimmune signaling in the hindbrain to obesity often is overlooked. This is a baffling oversight, as the DVC is important in both short-term and long-term control of body weight. The interconnected structures that comprise the DVC confer responsiveness to 'real-time' changes that occur during feeding, with pre-absorptive signals pertaining to gastrointestinal distension and nutrients in the gastrointestinal tract that may limit subsequent feeding transmitted by vagal afferents to the nucleus of the solitary tract (NTS)27. On the other hand, post-absorptive signals (e.g., circulating levels of metabolites or metabolic hormones), as well as hormonal signals of longer-term energy balance that may alter feeding or metabolic rate are monitored by the area postrema (AP), the hindbrain circumventricular organ. AP is uniquely situated to serve in this role by virtue of an incomplete blood-brain-barrier and receptors for numerous hormones28,29, including insulin and leptin12,13. Together, these structures detect and process signals related to both feeding and energy balance and then communicate the information directly and indirectly to other CNS areas involved in control of feeding and body weight, including hypothalamic areas30-32. Thus, decreases in TLR4 and GFAP in the DVC of rats that ran beginning three weeks after ovariectomy may be related to the detection of peripheral signals concomitant with the transient reduction in the rate of weight gain and, in turn, to communication within CNS pathways involved in the control of body weight.
Astrocytes influence communication in the CNS in multiple ways, including synthesis of glutamate and gamma-aminobutyric acid (GABA), clearance of potassium from the synapse, and long-term changes in synaptic efficacy33-35. A function of astrocytes that is particularly relevant to the DVC is their contribution to the blood-brain-barrier36,37. Thus, decreased GFAP in the DVC may indicate retraction of glial processes that contribute to maintenance of the blood-brain-barriere.g.,38 in the AP, thereby facilitating access of circulating hormones to receptors located in that region, and allowing consistent activation of leptin and/or insulin receptors despite reduced hormone levels—which may be critical to maintain neural and neuroimmune activation in CNS areas involved in body weight regulation. The idea of a neural-glial communication circuit in the hypothalamus has been proposed and is suggested to be related to innate immune responses39. Given the connectivity of the DVC16,30,31,40-42, it seems likely that this circuit includes the DVC. Thus, it is possible that the decrease in TLR4 levels in the DVC is an integral part of information flow within that circuit and is important to maintain activation in other CNS areas. Clearly, more studies are required to determine whether there are changes in the DVC blood-brain-barrier integrity as suggested by decreased GFAP levels, as well as the role of TLR4 activation in this communication circuit that affects feeding and body weight regulation; however, these are testable ideas. Thus, while still speculative, these observations could, at least in part, explain the recovery of weight gain after the first week of running, as well as the absence of differences in other neuroimmune factors.
Nonetheless, given the group differences in circulating levels of both leptin and insulin, it was surprising that these were the only changes observed. It is possible that changes in GFAP and TLR4 in the ARC and PVN-as well as changes in MCP-1 and IL-6 in any of the areas-are less likely with the overall slowing of weight gain during the last three weeks of the study. However, in our previous study, we found that neuroimmune factors in the ARC, PVN, and DVC change during post-ovariectomy weight gain as early as three weeks after ovariectomy10, arguing against this possibility. Other explanations merit further investigation. For example, the increase in body weight that occurred in both groups during the first three weeks may have 'set' the levels of neuroimmune factors such that more substantial weight loss is necessary to reverse those changes; residual estrogens during the first few weeks after OVX may have minimized changes in rats that run immediately after OVX; the transient nature of the exercise-induced slowing in weight gain-and the reduction in GFAP and TLR4 levels in the DVC-may have prevented changes in other neuroimmune factors. In short, although we cannot rule out the possibility that decreases in GFAP and TLR4 expression may be causally related to the slower rate of weight gain in rats with delayed access to running wheels, a more complete time-course study will be necessary to address the issues that remain, with additional studies to determine whether more substantial or more prolonged weight loss reveals differences in other neuroimmune factors.
Conclusion
Voluntary exercise had transient effects to slow the rate of weight gain in OVX rats, especially when exercise was initiated three weeks after ovariectomy. These transient effects on body weight were associated with decreased plasma leptin and insulin, and decreased levels of GFAP and TLR4 in the DVC. The latter may promote continued activation of receptors for insulin and/or leptin in the AP despite reduced circulating levels of insulin and leptin, and thereby sustain communication between CNS areas involved with the control of feeding and body weight regulation. As a result, body weight may be maintained even when running activity increases. In short, although the beneficial effects of exercise are increasingly emphasized as the obesity epidemic continues1, exercise may be accompanied by changes in metabolic hormones and neuroimmune factors that complicate its exclusive use as a weight-loss strategy. Thus, the possibility that reproductive hormones alter mechanisms associated with body weight regulation remains a challenge in understanding obesity and in identifying effective management strategies.
Acknowledgements
The authors gratefully acknowledge Daniel J. Buck, B.S. and Kelly McCracken, M.S. for technical assistance.
References
1. Levi J, Segal LM, St. Laurent R, Rayburn, J. The State of Obesity. Trust for America's Health and the Robert Wood Johnson Foundation. 2014. rwjf414829.pdf
2. Hales CM, Carroll MD, Fryar CD, Ogden DL. Prevalence of Obesity Among Adults and Youth: United States, 2015-2016. NCHS Data Brief; Hyattsville, MD: National Center for Health Statistics. 2017:288:1-8. https://www.cdc.gov/nchs/data/databriefs/db288.pdf
3. Managing Overweight and Obesity in Adults: Systematic Evidence Review From the Obesity Expert Panel. U.S. Department of Health and Human Services; National Institutes of Health; National Heart, Lung, and Blood Institute. 2013. http://www/nhlbi.nib.gov/guidelines
4. Cawley J and Meyerhoefer C. The Medical Care Costs of Obesity: An Instrumental Variables Approach. J Health Econ. 2012:31(1):219-230. https://doi.org/10.1016/j.jhealeco.2011.10.003
5. Kraschnewski JL, Boan J, Esposito J, Sherwood NE, Lehman EB, Kephart DK, Sciamanna CN. Long-term weight loss maintenance in the United States. Int J Obes. 2010:34(11):1644-1654. https://doi.org/10.1038/ijo.2010.94
6. Fothergill E, Guo J, Howard L, Kerns JC, Knuth ND, Brychta R, Chen KY, Skarulis MC, Walter M, Walter PJ, Hall KD. Persistent metabolic adaptation 6 years after "The Biggest Loser" competition. Obesity. 2016:8:1612-1619. https://doi.org/10.1002/oby.21538
7. Lovejoy JC, Sainsbury A. Sex differences in obesity and the regulation of energy homeostasis. Obesity Rev. 2009:10:154-167. https://doi.org/10.1111/j.1467-789X.2008.00529.x
8. Geary N, Asarian L. Cyclic estradiol treatment normalizes body weight and test meal size in ovariectomized rats. Physiol Behav. 1999:67(1):141-147. https://doi.org/10.1016/S0031-9384(99)00060-8
9. Graves NS, Hayes H, Fan L, Curtis, KS. Time course of behavioral, physiological, and morphological changes after estradiol treatment of ovariectomized rats. Physiol Behav. 2011:103:261-267. https://doi.org/10.1016/j.physbeh.2011.02.017
10. Curtis KS, McCracken K, Espinosa E, Ong J, Buck DJ, Davis RL. Temporal and site-specific changes in central neuroimmune factors during rapid weight gain after ovariectomy in rats. Neurochem Res. 2018:43(9):1802-1813. https://doi.org/10.1007/s11064-018-2596-6
11. Asarian L, Geary N. Sex differences in the physiology of eating. Am J Physiol Regul Integr Comp Physiol. 2013:305(11): R1215-R1267. https://doi.org/10.1152/ajpregu.00446.2012
12. Liberini, CG, Boyle CN, Cifani C, Venniro M, Hope BT, Lutz TA. Amylin receptor components and the leptin receptor are co-expressed in single rat area postrema neurons. Eur J Neurosci. 2016:43(5):653-661. https://doi.org/10.1111/ejn.13163
13. Unger JW, Moss AM, Livingston JN. Immunohistochemical localization of insulin receptors and phosphotryosine in the brainstem of the adult rat. Neurosci. 1991:42(3):853-861. https://doi.org/10.1016/0306-4522(91)90049-T
14. Morton GJ, Cummings DE, Baskin DG, Barsh GS, Schwarts MW. Central nervous system control of food intake and body weight. Nature Rev. 2006:443(21):289-295. https://doi.org/10.1038/nature05026
15. Woods SC, D'Alessio DA. Central control of body weight and appetite. J Clin Endocrinol Metab. 2008:93(11 Suppl 1):S37-50. https://doi.org/10.1210/jc.2008-1630.
16. Sohn J-W, Elmquist JK, Williams KW. Neuronal circuits that regulate feeding behavior and metabolism. Trends Neurosci. 2013:36:504-512. https://doi.org/10.1016/j.tins.2013.05.003
17. Kälin S, Heppner FL, Bechmann I, Prinz M, Tsch Öp MH, Yi C-X. Hypothalamic innate immune reaction in obesity Nat Rev Endocrinol. 2015:11:339-351. https://doi.org/10.1038/nrendo.2015.48
18. Valdearcos M, Xu AW, Koliwad SK. Hypothalamic Inflammation in the Control of Metabolic Function. Annu Rev Physiol. 2015:77:131-60. https://doi.org/10.1146/annurev-physiol-021014-071656
19. Rivera-Brown AM, WR Frontera. Principles of exercise physiology: responses to acute exercise and long-term adaptations to training. Physical Med Rehab. 2012:4:797-804. https://doi.org/10.1016/j.pmrj.2012.10.007
20. Moran TH. Gut peptide signaling in the controls of food intake. Obesity. 2006:14(Suppl 5):250S-253S. https://doi.org/10.1038/oby.2006.318
21. King JA, Garnham JO, Jackson AP, Kelly BM, Xenophontos S, Nimmo MA. Appetitie-regulatory hormone responses on the day following a prolonged bout of moderate intensity exercise. Physiol Behav 2015: 141: 23-31.
22. Maffei M, Halaas J, Ravussin E, Pratley RE, Lee GH, Zhang Y, Fei H, Kim S, Lallone R, Ranganathan S, Kern PA, Friedman JM. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat Med 1995:1:1155–1161. https://doi.org/10.1038/nm1195-1155
23. Swoap SJ, Bingaman MJ, Hult EM, Sandstrom NJ. Alternate-day feeding leads to improved glucose regulation on fasting days without significant weight loss in genetically obese mice. Am J Physiol Regul Integr Comp Physiol. 2019:317:R461-R469. https://doi.org/10.1152/ajpregu.00140.2019
24. Eckel LA, Houpt TA, Geary N. Spontaneous meal patterns in female rats with and without access to running wheels. Physiol Behav. 2000:70: 397-405. https://doi.org/10.1016/S0031-9384(00)00278-X
25. Ryan KK, Woods SC, Seely RJ. Central nervous system mechanisms linking the consumption of palatable high-fat diets to the defense of greater adiposity. Cell Metab. 2012: 15:137-149.
26. Dalby MJ, Aviello G, Ross AW, Walker AW, Parrett P, Morgan PJ. Diet induced obesity is independent of metabolic endotoxemia and TLR4 signalling, but markedly increases hypothalamic expression of the acute phase protein, SerpinA3N. Sci Reports. 2018: 8:15648
27. Iwasaki Y, Yada T. Vagal afferents sense meal-associated gastrointestinal and pancreatic hormones: Mechanism and physiological role. Neuropeptides. 2012:46:291-297. https://doi.org/10.1016/j.npep.2012.08.009
28. Price CJ, Hoyda TD, Ferguson AV. The Area Postrema: A brain monitor and integrator of systemic autonomic state. Neuroscientist. 2008: 14(2):182-194. https://doi.org/10.1177/nro.2008.
29. Cabral A, Cornejo, MP, Fernandez G, De Francesco PN, Garcia-Romero G, Uriarte M, Zigman JM, Portiansky E, Reynaldo M, Perello M. Circulating ghrelin acts on GABA neurons of the area postrema and mediates gastric emptying in male mice. Endocrinol. 2017:158:1436-1449. https://dio.org/10.1210/en.2016-1815.
30. Ter Horst GJ, de Boer P, Luiten PG, van Willigen JD. Ascending projections from the solitary tract nucleus to the hypothalamus. A Phaseolus vulgaris lectin tracing study in the rat. Neurosci. 1989:31:785-797. https://doi.org/ns10.1016/0306-4522(89)90441-7
31. Utoyama M, Akieda-Asai S, Koda S, Nunoi H, Date Y. Role of the neural pathway from hindbrain to hypothalamus in the regulation of energy homeostasis in rats. Neurosci Lett. 2016:614:83-88. https://doi.org/10.1016/j.neulet.2016.01.005
32. Cunningham ET, Miselis RR, Sawchenko PE. The relationship of efferent projections from the area postrema to vagal motor and brain stem catecholamine-containing cell groups: an axonal transport and immunohistochemical study in the rat. Neurosci. 1994:58:635-648. https://doi.org/ns10.1016/0306-4522(94)90087-6
33. Allen NJ, Barres BA. Signaling between glia and neurons: focus on synaptic plasticity. Current Opinion Neurobio. 2005:15(5):542-548. https://doi.org/10.1016/j.conb.2005.08.006
34. Barres BA. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 2008:60(3):430-440. https://doi.org/10.1016/j.neuron.2008.10.013
35. Halassa MM, Haydon PG. Integrated brain circuits: Astrocytic networks modulate neuronal activity and behavior. Ann Rev Physiol. 2010:72:335-355. https://doi.org/10.1146/annurev-physiol-021909-135843
36. Liebner S, Czupalla CJ, Wolburg H. Current concepts of blood-brain barrier development. Int. J. Dev. Biol. 2011:55:467-476. https://doi.org/10.1387/ijdb.103224sl
37. Sun M, McDonald SJ, Brady RD, Collins-Praino L, Yamakawa GR, Monif M, O'Brien TJ, Cloud GC, Sobey CG, Mychasiuk R, Loane DJ, Shuttz SR. The need to incorporate aged animals into the preclinical modeling of neurological conditions. Neurosci Biobehav Rev. 2020: 109: 114-128.
38. Hsuchou H, Kastin AJ, Mishra PK, Pan W C-Reactive Protein Increases BBB Permeability: Implications for Obesity and Neuroinflammation. Cell Physiol Biochem 2012:30:1109-1119
39. Kalin S, Heppner FL, Bechmann I, Prinz M, Tschop MH, Yi C-X. Hypothalamic innate immune reaction in obesity. Nat Rev Endocrinol. 2015: 11: 339-351.
40. Hosoya Y, Matsushita M. A direct projection from the hypothalamus to the area postrema in the rat, as demonstrated by the HRP and autoradiographic methods. Brain Res. 1981:214:144-149. https://doi.org/10.1016/0006-8993(81)90445-5
41. Sawchenko PE, Swanson LW. Immunohistochemical identification of neurons in the paraventricular nucleus of the hypothalamus that project to the medulla or to the spinal cord in the rat. J Comp Neurol. 1982:205:260-272. https://doi.org/10.1002/cne.902050306
42. Kirchgessner AL, Sclafani A, Nilaver G. Histochemical identification of a PVN-hindbrain feeding pathway. Physiol Behav. 1988:42:529-543. https://doi.org/10.1016/0031-9384(88)90154-0
Figure Legends
Figure 1.Body weight in ovariectomized (OVX) rats initially given access to running wheels for three weeks and then switched to standard cages (R-S) or initially housed in standard cages for three weeks before being given access to running wheels for three weeks (S-R).
Panel A: body weight for rats in the R-S group (open squares) and for rats in the S-R group (black circles). * = significantly greater than preceding weeks within the R-S group (ps<0.05-0.001); # = significantly greater than preceding weeks within the S-R group (ps<0.05-0.001); @ = significantly greater than week 4 within S-R group (p<0.01)
Panel B: weekly change in body weight for rats in the R-S group (open bars) and for rats in the S-R group (black bars). P-values (0.01, 0.001) indicate significant difference between groups at specific time points; white # = significantly less than all other time points for both groups (ps<0.05-0.001); * = significantly less than change from week 1-2, 2-3 (ps<0.05-0.01) within R-S group; black # = significantly less than change from week 0-1, 1-2, 2-3 (ps<0.001) within S-R group
Figure 2. Weekly distance run by ovariectomized (OVX) rats initially given access to running wheels for three weeks and then switched to standard cages (R-S; open squares) or initially housed in standard cages for three weeks before being given access to running wheels for three weeks (S-R; black circles). * = significantly less than other weeks, independent of group (ps<0.05, 0.001)
Figure 3. Plasma levels of insulin (Panel A) and Leptin (Panel B) in ovariectomized (OVX) rats initially given access to running wheels for three weeks and then switched to standard cages (R-S; open bars) or initially housed in standard cages for three weeks before being given access to running wheels for three weeks (S-R; black bars). Brackets and p-values (0.05) indicate significant differences between groups
Figure 4. Neuroimmune factors in central nervous system areas involved in body weight regulation in ovariectomized (OVX) rats initially given access to running wheels for three weeks and then switched to standard cages (R-S; open bars) or initially housed in standard cages for three weeks before being given access to running wheels for three weeks (S-R; black bars).
Panel A: Monocyte chemoattractive protein-1 (MCP-1) in the hypothalamic arcuate nucleus (ARC) and paraventricular nucleus (PVN) and in the hindbrain dorsal vagal complex (DVC).
Panel B: Interleukin-6 (IL-6) in the hypothalamic ARC and PVN and in the hindbrain DVC.
Panel C: Glial fibrillary acidic protein (GFAP) in the hypothalamic ARC and PVN and in the hindbrain DVC. Brackets and p-value (0.05) indicate significant difference between groups within an area
Panel D: Toll-like receptor-4 (TLR4) in the hypothalamic ARC and PVN and in the hindbrain DVC. Brackets and p-value (0.05) indicate significant difference between groups within an area
Panel E: Representative digital images of western blots for TLR4 (top), the housekeeping protein β-tubulin (middle), and GFAP in the DVC. MW 'ladder' is in the first lane, other lanes are from individual S-R and R-S rats. Images were adjusted for brightness and contrast.
Table 1. Spleen weight and plasma protein concentrations in ovariectomized (OVX) rats initially given access to running wheels for three weeks and then switched to standard cages (R-S) or initially housed in standard cages for three weeks before being given access to running wheels for three weeks (S-R). * = significantly less than other group (p<0.05)
|
R-S |
S-R |
| Spleen (mg/100g body wt) |
187.1 ± 8.0 |
211.3 ± 9.9 |
| Plasma proteins (g/dL) |
7.61 ± 0.16 |
7.18 ± 0.07* |