Avery Dutcher., D.O., Oklahoma State University Center for Health Sciences, Tulsa OK
Andriana Pena, D.O., Oklahoma State University Center for Health Sciences, Tulsa Ok
Liming Fan, M.S., Oklahoma State University Center for Health Sciences, Tulsa OK
Al Rouch., Ph.D., Oklahoma State University Center for Health Sciences, Tulsa OK
Abstract
Background: Sex differences in renal function are well known. The purpose of this study was to determine if sex differences in urinary protein excretion (PE) occur in mice consuming a high protein diet and to investigate the potential roles of the sex steroids estrogen (E2) and testosterone (T).
Methods: Four experiments each with the same procedure were conducted. The first included intact male (IM) and castrated (CAS) male mice (n=6/group), second included intact female (IF) and ovariectomized (OVX) female mice (n=6/group), third included IF, OVX treated with placebo (P), and OVX treated with estrogen (E2) mice (n=4/group), and fourth included IM, CAS treated with P, and CAS treated with T mice (n=4/group). All mice were placed in metabolic cages and consumed a high protein diet for 25 days. PE (mg/day) was determined from the measurement of total protein in a 24-hr urine collection from each mouse. PE was measured on days 2, 7, 14, 21, and 25 and values were averaged per group.
Results: When averaged for 25 days, PE for IM mice was higher than for CAS mice (20.8 ± 2.2 vs 5.4 ± 1.2, respectively, p < 0.001). PE was not different between IF and OVX mice (7.7 ± 1.1 vs 5.3 ± 2.3, respectively). PE was not different between IF, OVX-P, and OVX-E2 mice (7.1 ± 2.5 vs 5.2 ± 1.0 vs 6.5 ± 1.0, respectively). PE for IM mice was not different than for CAS-T mice (18.2 ± 2.9 vs 20.7 ± 2.2, respectively) and both were higher than CAS-P mice (5.1 ± 1.0, p < 0.001).
Conclusions: Testosterone induces proteinuria in mice consuming high protein whereas E2 plays no role or only a minor role in proteinuria under these experimental conditions. Our results suggest that androgens may account for the higher incidence of kidney disease in males compared to age-matched, premenopausal females.
Keywords: sex differences, proteinuria, testosterone, estrogen, high protein diet
Introduction
The obesity epidemic is a global health concern and presents significant challenges for the medical profession. While many factors contribute to this growing epidemic, personal life-style choices related to physical activity and dietary manipulation undoubtedly are key to reducing the incidence of obesity 1. Major comorbidities associated with obesity involve pathologies of the cardiovascular and renal systems. Interestingly, research has demonstrated the effectiveness of high protein (HP) diets on weight reduction 2-4. Consequently, diets known for low carbohydrate and elevated protein, such as the Atkins and Keto diets, have become quite popular.
While HP ingestion might prove beneficial with weight reduction, it also increases the workload of the renal system 3,5,6. The body does not store elevated ingested protein as occurs with elevated lipid and carbohydrate ingestion. Metabolism of ingested protein, primarily in the liver, results in products destined for renal excretion, particularly urea and amino acids. In this process glomerular filtration and the urinary concentrating mechanism increase in activity thus increasing the metabolic work of the kidney. High protein consumption results in kidney hypertrophy, and potential renal injury 7-12. It should be noted that the source of dietary protein might be important with respect to specific kidney function and potential pathologies. For example, soy and whey sources might be preferable compared to casein and animal protein13-15.
Biological men have a higher incidence of renal disease than age-matched, premenopausal biological women and moreover, when controlling for all other risk factors, male biology is associated with faster progression of renal disease 6. Interestingly, postmenopausal biological females also appear to lose their renoprotection 16,17. Relationships between the progression of renal disease and the presence of testosterone, or the absence of estrogen, or the altered ratio of androgen-to-estrogen remain undetermined.
As the ingestion of HP diets and the impact of sex hormones may independently enhance the progression of renal disease, it becomes important to study these two factors in combination especially now with more people ingesting HP diets to combat obesity. The purpose of this study was to determine if biological sex differences impact proteinuria in mice consuming a HP diet. In addition, we investigated the role of the major sex steroid hormones estrogen and testosterone on these differences. Based on previous studies regarding the renal excretion of total protein in rodents 18-20 and the current consensus that estrogen protects against renal disease while testosterone promotes opposite effects 21-31, we hypothesized that estrogen replacement in ovariectomized female mice and testosterone replacement in castrated male mice would decrease and increase protein excretion (PE), respectively.
Methods
This study was designed to investigate potential sex differences, the potential roles of estrogen and testosterone, and impact of increased dietary protein ingestion on the rates of renal PE in mice. This study was conducted to answer the following questions: 1) do sex differences regarding PE exist in mice consuming a HP diet; 2) does estrogen play a role in preventing excessive PE; and 3) does testosterone play a role in promoting excessive PE.
Male and female mice (Hsd:ICR (CD-1®)) at 30-35 day old and 20-30 g body weight were purchased from Envigo, Inc. (Indianapolis, IN). Envigo technicians performed the ovariectomy and castration 10-14 days prior to the studies. Upon arrival to the institution’s animal facilities, mice were placed in plastic cages under standard conditions for 4-7 days with standard rodent chow (Harlan Teklad 18% protein) and water before being placed in metabolic cages. The Oklahoma State University Center for Health Sciences Institutional Animal Care and Use Committee approved all experimental procedures with animals.
Experiments were conducted with mice in metabolic cages. Daily measurements included body weight, food and water intake, urine volume (UV), and urine protein level via dipstick. Total protein concentration in the urine was measured at the OSU-CHS Medical Center via a colorimetric method using an Olympus AU400TM analyzer. Renal protein excretion (PE, mg/day) was determined by multiplying the total protein concentration (mg/ml) in the urine by the UV (ml/day). Upon entering metabolic cages, all mice consumed a high - 40% protein diet (Teklad Custom Diet, Envigo, TD.90018) with water ad libitum throughout the experimental protocols. Constituents (g/Kg) of this diet included casein (460), sucrose (232), corn starch (200), corn oil (50), cellulose (15), calcium phosphate (9.6), calcium carbonate 10.2, and a vitamin mix (10).
Four experiments were conducted. The first included 6 intact males (IM) and 6 castrated males (CAS). The second included 6 intact females (IF) and 6 ovariectomized females (OVX). The third included 4 IF, 4 OVX mice treated with P, and 4 OVX mice treated with exogenous estrogen (E2). The fourth included 4 IM, 4 CAS mice treated with P, and 4 CAS mice treated with exogenous testosterone (T). Experiments lasted 25 days and PE was measured on days 2, 7, 14, 21, and 25. Treatments involved specialized pellets (placebo, 0.7 mg E2, or 25 mg testosterone) implanted subcutaneously to provide continuous hormone release producing physiological levels of E2 and T. (Innovative Research of America, Sarasota, FL). At the end of each experiment mice were sacrificed via isoflurane anesthesia followed by cervical dislocation. Kidneys were removed and weighed to determine kidney-to-body weight.
Statistics: Repeated measures ANOVA using Graphpad Prism 9.0 were used to determine differences in PE by measuring the mean PE of each group at each time point. One-way ANOVA was used for comparisons of kidney-to-body weight differences between groups.
Results
Figure 1 shows the average daily food intake of mice from the four experiments. Average food intake and thus protein intake did not differ between groups in any experiment.
Figure 1. Food intake was not different between groups of mice in any experiment
Figure 1. Average daily food intake of each experiment: A) IF and OVX mice, B) IM and CAS mice, C) IF, OVX-P, and OVX-E2 mice, and D) IM, CAS-P, and CAS-T mice. No differences in food intake were measured between groups in any experiment. Analysis conducted via one-way ANOVA.
Figure 2 shows PE data from each experiment. Experiment 1: IF mice showed non-significantly higher PE than OVX mice except in week 3 where IF PE was significantly higher. Experiment 2: IM mice showed significantly higher PE than CAS mice. Experiment 3: No differences in PE occurred between IF, OVX-P, and OVX-E2 mice. Experiment 4: IM mice and CAS-T mice showed similarly high PE which were significantly higher than CAS-P mice.
Figure 2. IM mice show higher PE than CAS mice and exogenous testosterone increases PE in CAS mice under high protein ingestion
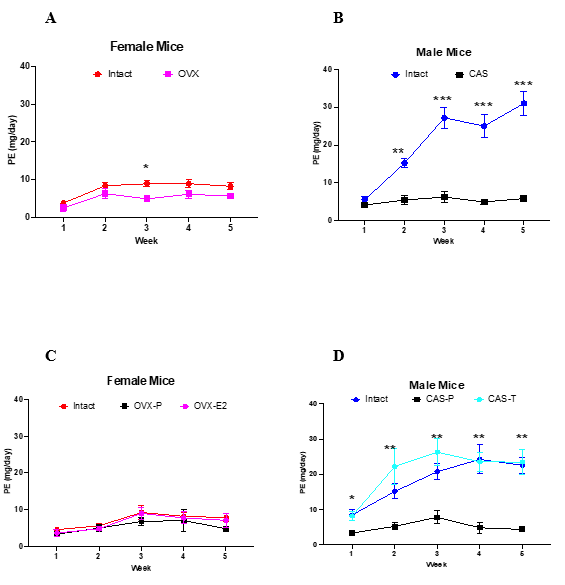
Figure 2. Renal PE (mg/day, mean ± se) of each week from each experiment. A) IF mice had non-significantly higher PE than OVX mice except in the third week where IF mice had significantly higher PE (*p<0.05). B) IM mice had significantly higher PE compared to CAS mice throughout the study, (**p<0.01, ***p<0.001). C) No differences in PE were measured between IF, OVX-P, or OVX-E2 mice. D) IM and CAS-T mice had significantly higher PE than CAS-P mice throughout the study. (*p<0.05, **p<0.01). Data analyzed via repeated measures ANOVA.
Table 1. PE (mg/day, mean ± se) from each week of each experiment
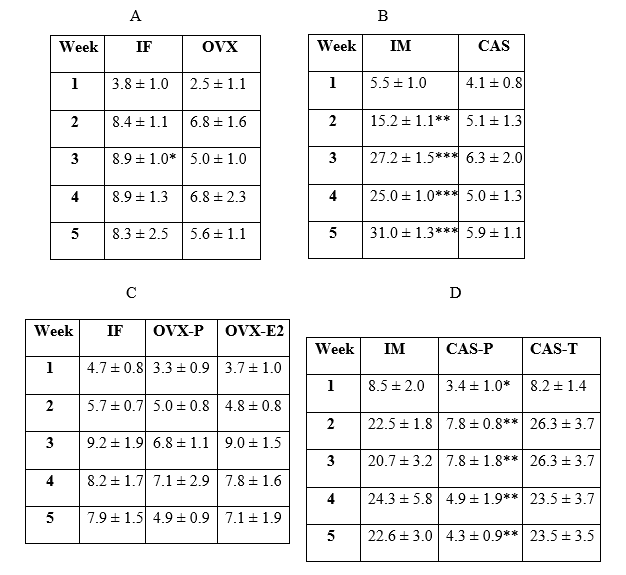
Table provides mean ± se of PE (mg/day) results from each weekly measurement period of each experiment shown in figure 2 (Table 1A – Figure 2A; Table 1B – Figure 2B; Table 1C – Figure 2C, and Table 1D – Figure 2D). (Table 1A: IF>OVX, *p<0.05), (Table 1B: IM>CAS, **p<0.01 & ***p<0.001), (Table 1D: IM & CAS-T > CAS-P, *p<0.05 & **p<0.01) All significant differences were determined via repeated measures ANOVA.
Figure 3 shows the ratio of kidney weight-to-body weight of each group from experiments 3 and 4. CAS-P mice had the lowest values which were not different from those of the female groups where no differences were measured. CAS-T mice had the highest values.
Figure 3. Kidney-to-Body Weight Ratio in Male and Female Mice
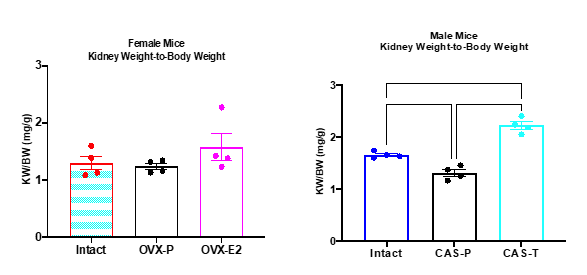
Figure 3. Ratio of kidney weight-to-body weight. No differences were measured in female mice. IM mice had higher values than CAS+P mice (p<0.010) and CAS+T mice had higher values than IM (p<0.001) and CAS+P mice (p<0.0001). Analyzed via one-way ANOVA.
Discussion
Biological males have a higher incidence of renal disease than age-matched premenopausal biological females and diseased kidneys in both sexes worsen progressively faster in biological males 19,32-34. The underlying mechanisms of these sex differences and pathological effects are not completely understood, but it is widely accepted that sex steroids have important roles. Renoprotection in premenopausal biological females suggests that estrogen provides protection and/or testosterone induces potential harm.
Excessive proteinuria is one of the major indicators of kidney disease. Normal kidneys prevent most protein from filtering through the glomerulus and that which does filter through gets metabolized by the nephron 35. Thus, only very small amounts of protein normally appear in the urine. Diets high in protein and low in carbohydrate have become popular in combating obesity. Elevated protein ingestion changes renal function particularly with respect to increased glomerular filtration and urinary concentration 6,7,11.
Our first experiment showed that IF mice had slightly higher, though for the most part, not significantly different PE compared to OVX mice. The most obvious difference in PE resulted from experiments with male mice. IM mice demonstrated significantly higher PE compared to CAS mice. Comparison of PE between CAS mice with either the IF or OVX mice showed no significant differences.
Results from the first two experiments suggested that testosterone plays a role in PE in male mice but did not provide much support for any role of estrogen in female mice. We conducted two more experiments where we included intact, gonadectomized, and gonadectomized mice with exogenous E2 and T administration in females and males, respectively. The administration of E2 to OVX mice had no effect on PE when compared to the IF and OVX-P mice. However, administration of T to CAS mice resulted in high PE values equal to those of IM mice.
It has long been known that males have larger kidneys than females and the androgens play the key role in kidney size 36. Moreover, high protein ingestion promotes kidney hypertrophy probably due to the increased metabolic activity imposed on the kidney 8,12,36-38. Hypertrophy of cells or organs often is a prelude to a pathological event. Of particular clinical importance comes from a study by Schrijvers et al. where high protein intake in mice increases glomerular hypertrophy and perhaps results in glomerular injury 11. While we did not separate the effects of HP intake and testosterone on kidney size, our results shown in Figure 3 indicate that kidney hypertrophy could be of major concern in males consuming a HP diet. The combination of HP and testosterone enhances the risk for detrimental kidney hypertrophy. Kidneys from CAS and CAS-P mice were the same size as female kidneys.
We acknowledge weaknesses of the current study. We measured total protein in the urine and did not identify specific protein sources. We suspect that at least some of this protein is albumin, which under normal conditions should not appear in the urine. Dipstick results from this study (data not shown) indicate similar findings as the measured PE. This study did not begin and end with a normal protein diet but rather tested mice ingesting only the HP diet. We also did not measure circulating levels of E2 and T in mice. Previously, we measured E2 levels in gonadectomized female mice with the same E2 pellet used in the current study and results indicated values in the physiological range 39. As stated above, this study did not determine the effect of HP alone on kidney hypertrophy. Future studies are required to correct these deficiencies.
Conclusion
In summary, our goal was to determine if sex differences occur in renal PE in mice consuming a HP diet and the potential roles of E2 and T. We conclude: 1) sex differences in renal PE indeed exist with male mice having much higher PE than female mice; 2) estrogen had no effect on PE; 3) testosterone increased PE. This study expands overall knowledge on sex differences in renal function under specific conditions of HP diets. Further research in the role of testosterone on PE should determine the specific mechanisms of how this sex steroid increases PE as well as any long-term potential detrimental effects would add to this body of knowledge. While additional research needs to be performed, these results have important implications for those considering a long-term dietary regimen of high protein.
REFERENCES
1. T.A. W, Tronieri JS, Butryn ML. Lifestyle modification approaches for the treatment of obesity in adults. Am Psychol. 2020;75(2):235-251. doi:10.1037/amp0000517
2. Astrup A, Raben A, Geiker N. The role of higher protein diets in weight control and obesity-related comorbidities. Int J Obes (Lond). May 2015;39(5):721-6. doi:10.1038/ijo.2014.216
3. Eisenstein J, Roberts SB, Dallal G, Saltzman E. High-protein weight-loss diets: Are they safe and do they work? A review of the experimental and epidemiologic data. Nutrition Reviews. 2002;60(7):189-200.
4. Lacroix M, Gaudichon C, Martin A, et al. A long-term high-protein diet markedly reduces adipose tissue without major side effects in Wistar male rats. Am J Physiol Regul Integr Comp Physiol. 2004;287:R934-R942.
5. Bouby N, Bankir L. Effect of high protein intake on sodium, potassium-dependent adenosine triphosphatease activity in the thick ascending limb of Henle's loop in the rat. Clin Sci. 1988;74:319-329.
6. Brenner BM, Meyer TW, Hostetter TH. Dietary protein intake and the progressive nature of kidney disease: the role of hemodynamically mediated glomerular injury in the pathogenesis of progressive glomerular sclerosis in aging, renal ablation, and intrinsic renal disease. N Engl J Med. Sep 9 1982;307(11):652-9. doi:10.1056/NEJM198209093071104
7. Benabe JE, Cordova HR. Effect of high-protein diet on renal concentration capacity in rabbits. Am J Physiol. 1992;263 (Renal Fluid Electrolyte Physiol. 32):F1051-F1056.
8. Chin E, Bondy CA. Dietary protein-induced renal growth: correlation between renal IGF-1 synthesis and hyperplasia. Am J Physiol Cell Physiol. 1994;266:C1037-C1045.
9. Hammond KA, Janes DN. The effects of increased protein intake on kidney function. J Exp Biol. 1998;201:2081-2090.
10. Hostetter TH, Meyer TW, Rennke HG, Brenner BM. Chronic effects of dietary protein in the rat with intact and reduced renal mass. Kidney International. 1986;30:509-517.
11. Schrijvers BF, Rasch R, Tilton RG, Flyvbjerg A. High protein-induced glomerular hypertrophy is vascular endothelial growth factor-dependent. Kidney International. 2002;61:1600-1604.
12. van Neck JW, Cingel V, van Vliet AK, Drop SLS, Flyvbjerg A. High-protein induced renal enlargement is growth hormone independent. Kidney International. 2002;62:1187-1195.
13. Hall WL, Millward DJ, Long SJ, Morgan LM. Casein and whey exert different effects on plasma amino acid profiles, gastrointestinal hormone secretion and appetite. Br J Nutr. 2003;89:239-248.
14. Nevala R, Vaskonen T, Vehniainen J, Korpela R, Vapaatalo H. Soy based diet attenuates the development of hypertension when compared to casein based diet in spontaneously hypertensive rat. Life Sci. 2000;66(2):115-24.
15. Song S, Hooiveld GJ, Li M, et al. Dietary soy and meat proteins induce distinct physiological and gene expression changes in rats. Sci Reports. 2016;620036. doi:10.1038
16. Reckelhoff JF. Sex steroids, cardiovascular disease, and hypertension. Hypertension. 2005;45:170-174.
17. Reckelhoff JF, Yanes LL, Iliescu R, Fortepiani LA, Granger JP. Testosterone supplementation in aging men and women: possible impact on cardiovascular-renal disease. Am J Physiol Renal Physiol. 2005;289:F941-F948.
18. Baylis C. Age-dependent glomerular damage in the rat. J Clin Invest. 1994;94:1823-1829.
19. Reckelhoff JF, Zhang H, K. S. Gender differences in development of hypertension in spontaneously hypertensive rats. Hypertension. 2000;35(2):480-483.
20. Remuzzi A, Puntorieri S, Mazzoleni A, Remuzzi G. Sex related differences in glomerular ultrafiltration and proteinuria in Munich-Wistar rats. Kidney International. 1988;34:481-486.
21. Blush J, Lei J, Ju W, Silbiger SR, Pullman J, Neugarten J. Estradiol reverses renal injury in AIb/TGF-b1 transgenic mice. Kidney International. 2004;66:2148-2154.
22. Dixon A, Maric C. 17b-Estradiol attenuates diabetic kidney disease by regulating extracellular matrix and transforming growth factor-b protein expression and signaling. Am J Physiol Renal Physio. 2007;293:F1678-F1690.
23. Dubey RK, Jackson EK. Estrogen-induced cardiorenal protection: potential cellular, biochemical, and molecular mechanisms. Am J Physiol Renal Physiol. 2001;280:F365-F388.
24. Elliot SJ, Berho M, Korach K, et al. Gender-specific effects of endogenous testosterone: female a-estrogen receptor-deficient C57BI/6J mice develop glomerulosclerosis. Kidney International. 2007;72:464-472.
25. Elliot SJ, Karl M, Berho M, et al. Estrogen deficiency accelerates progression of glomerulosclerosis in susceptible mice. Am J Path. 2003;162(5):1441-1448.
26. Ji H, Menini S, Mok K, et al. Gonadal steroid regulation of renal injury in renal wrap hyhpertension. Am J Physiol Renal Physiol. 2004;288:F513-F520.
27. Karl M, Berho M, Pignac-Kobinger J, Striker GE, Elliot SJ. Differential effects of continuous and intermittent 17b-estradiol replacement and tamoxifen therapy on the prevention of glomerulosclerosis. Am J Path. 2006;169(2):351-361.
28. Mankhey RW, Wells CC, Bhatti F, Maric C. 17b-Estradiol supplementation reduces tubulointerstitial fibrosis by increasing MMP activity in the diabetic kidney. Am J Physiol Regul Integr Comp Physiol. 2007;292:R769-R777.
29. Maric C, Sandberg K, Hinojosa-Laborde C. Glomerulosclerosis and tubulointerstitial fibrosis are attenuated with 17b-estradiol in the aging Dahl salt sensitive rat. J Am Soc Nephrol. 2004;15:1546-1556.
30. Sun J, Devish K, Langer WJ, Carmines PK, Lane PH. Testosterone treatment promotes tubular damage in experimental diabetes in prepubertal rats. Am J Physiol Renal Physiol. 2007;292:F1681-F1690.
31. Verzola D, Gandolfo MT, Salvatore F, et al. Testosterone promotes apoptotic damage in human renal tubular cells. Kidney International. 2004;65:1252-1261.
32. Cattran DC, Reich HN, Beanlands HJ, Miller JA, Scholey JW, Troyanov S. The impact of sex in primary glomerulonephritis. Nephrol Dial Transplant. 2008;23:2247-2253.
33. Lane PH. Estrogen receptors in the kidney: lessons from genetically altered mice. Gender Medicine. 2008;5(Suppl. A):S11-S18.
34. Neugarten J, Acharya A, Silbiger SR. Effect of gender on the progression of nondiabetic renal disease: a meta-analysis. J Am Soc Nephrol. 2000;11:319-329.
35. Koeppen BM, Stanton BA. Renal Physiology. 6th ed. Elsevier; 2019.
36. Jean-Faucher C, Berger M, Gallon C, de Turckheim M, Veyssiere G, Jean C. Sex-related differences in renal size in mice: ontogeny and influence of neonatal androgens. J Endocrinol. Nov 1987;115(2):241-6.
37. Jackson Jr H, Riggs MD. The effect of high protein diets on the kidneys of rats. J Biol Chem. 1926;67(1):101-107.
38. Moise TL, Smith AH. The effect of high protein on the kidneys. An experimental study. Arch Path & Lab Med. 1927;4(4):530-542.
39. Rouch AJ, Meter J, Fan L. Effect of estrogen on blood pressure and renal sodium handling in mice consuming a high salt high fructose diet: A pilot study. OK Med Proc. 2018;2(2)